Domestiquée il y a 6 000 à 10 000 ans sur les hauteurs andines du Sud du Pérou [1, 2], la pomme de terre cultivée (Solanum tuberosum L.) est rapidement devenue une denrée alimentaire fondamentale dans le monde entier [3]. Avec une production mondiale de 377 millions de tonnes en 2016, pour une valeur brute totale de 111 milliards de dollars, la pomme de terre se hisse en quatrième position parmi les plantes cultivées dans le monde, derrière le riz, le blé et le maïs [4].
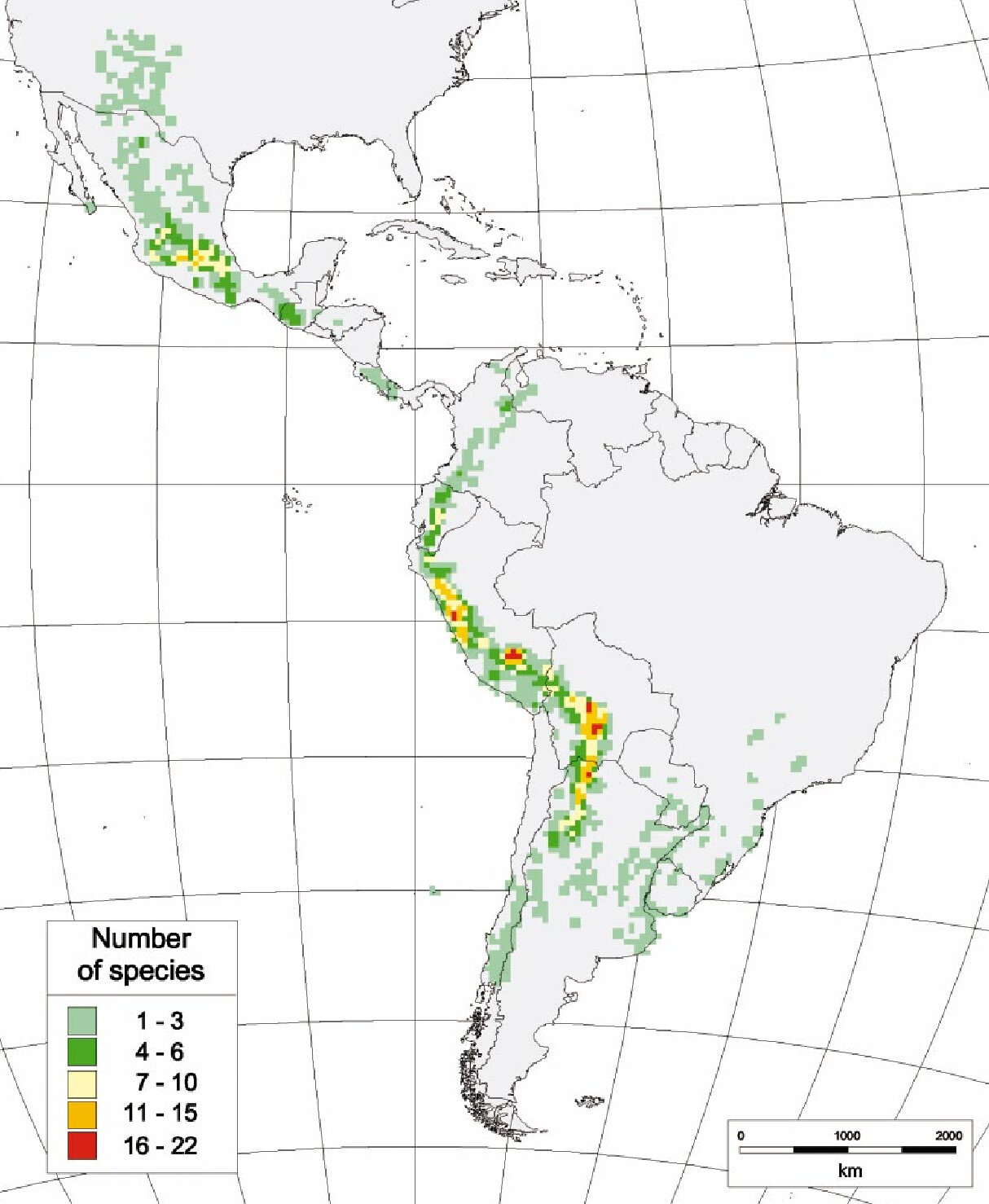
Les directions prises actuellement pour l’amélioration de la pomme de terre visent à augmenter les rendements, la compatibilité avec les besoins de l’industrie agroalimentaire, la résistance aux stress abiotiques (p. ex., la sécheresse) et la résistance aux pathogènes [5, 6]. Pour identifier des traits d’intérêt à introduire chez S. tuberosum, les améliorateurs peuvent tirer parti de la grande diversité botanique des pommes de terre, qui comprend 4 espèces cultivées et 107 espèces sauvages [2], ditribuées dans toute l’Amérique latine (Figure 1), du Nouveau-Mexique à la Patagonie [7, 8].
Adaptées à un large éventail de conditions environnementales et climatiques, les pommes de terre sauvages possèdent des traits d’intérêt pour l’agriculture, comme une plus grande résistance aux pathogènes et à la sécheresse. Plusieurs études génomiques effectuées récemment dans le clade des pommes de terre (Solanum sect. Petota) ont en effet mis au jour l’extraordinaire diversité génétique des espèces sauvages et l’utilité de celle-ci pour l’amélioration variétale [9, 10].
S. chacoense: une ressource pour l’amélioration de la pomme de terre…
Mon projet de recherche concerne la biologie reproductive de Solanum chacoense (Figure 2), une espèce sauvage de pomme de terre dont le génome a été récemment séquencé [11]. S. chacoense intéresse la recherche agronomique en raison de sa grande résistance à une vaste gamme de pathogènes, comprenant le virus Y de la pomme de terre [12, 13], les bactéries causant le flétrissement et la verticilliose [14, 15] ainsi que la gale commune [16, 17], ou encore les champignons responsables de la pourriture du collet et de la maladie de la jambe noire [18], et du mildiou [19].
Solanum chacoense possède également un métabolisme secondaire particulier, produisant de grandes quantités de glycoalcaloïdes particuliers, comme les leptines et les leptinines [20], qui sont particulièrement efficaces contre le doryphore de la pomme de terre [21]. Ce petit insecte herbivore (Figure 3) est un dangereux nuisible, qui peut détruire complètement une culture de pommes de terre en se nourissant des feuilles. Des travaux sont actuellement menés pour mieux comprendre les gènes impliqués dans ces voies métaboliques [22, 23] et pour les introduire chez la pomme de terre cultivée [24, 25, 26, 27].
…et une plante-modèle pour l’étude de la reproduction sexuée
Étant diploïde, S. chacoense est également utilisée comme modèle alternatif à la pomme de terre cultivée tétraploïde pour la biologie moléculaire et cellulaire, en particulier pour la recherche sur la reproduction des plantes. Par exemple, dans notre institut de recherche, S. chacoense a été utilisée au labo Cappadocia pour décrypter les mécanismes gouvernant l’auto-incompatibilité gamétophytique à S-RNase [28, 29, 30, 31]. Le labo Geitmann, désormais installé à l’Université McGill, a également employé S. chacoense dans ses recherches sur la cytomécanique du tube pollinique [32, 33, 34].
Le labo Matton a découvert plusieurs protéines impliquées dans des voies de signalisation reproductives chez S. chacoense, dont des peptides RALF [35, 36], des MAP kinases (missing reference) et des récepteurs kinases [37, 38, 39, 40]. Notre équipe a également produit des études transcriptomiques [41], protéomiques [42] et sécrétomiques [43] à large spectre pour mieux comprendre la reproduction de S. chacoense.
Mon projet de doctorat vise à comprendre comment les interactions pollen–pistil, en particulier le guidage du tube pollinique, sont impliquées dans l’isolement reproductif des pommes de terre sauvages, c’est-à-dire les mécanismes qui leur permettent d’éviter l’hybridation interspécifique.


Références
- Spooner DM, McLean K, Ramsay G, Waugh R and Bryan GJ. (2005). A single domestication for potato based on multilocus amplified fragment length polymorphism genotyping. Proc. Natl. Acad. Sci. U. S. A., 102(41): 14694–9. DOI: 10.1073/pnas.0507400102
- Spooner DM, Ghislain M, Simon R, Jansky SH and Gavrilenko T. (2014). Systematics, diversity, genetics, and evolution of wild and cultivated potatoes. Bot. Rev., 80(4): 283–383. DOI: 10.1007/s12229-014-9146-y
- Birch PRJ, Bryan G, Fenton B, Gilroy EM, Hein I, Jones JT, … Toth IK. (2012). Crops that feed the world 8: Potato: are the trends of increased global production sustainable? Food Sec., 4(4): 477–508. DOI: 10.1007/s12571-012-0220-1
- Food and of the United Nations AO. (2016). FAO Statistics Division data for year 2016. URL: http://www.fao.org/faostat/en (accessed August 1, 2018).
- Halterman D, Guenthner J, Collinge S, Butler N and Douches D. (2015). Biotech potatoes in the 21st century: 20 years since the first biotech potato. Am. J. Potato Res., 93(1): 1–20. DOI: 10.1007/s12230-015-9485-1
- Hameed A, Zaidi SS-E-A, Shakir S and Mansoor S. (2018). Applications of new breeding technologies for potato improvement. Front. Plant Sci., 9: 925. DOI: 10.3389/fpls.2018.00925
- Hijmans RJ and Spooner DM. (2001). Geographic distribution of wild potato species. Am. J. Bot., 88(11): 2101–12.
- Hijmans RJ, Spooner DM, Salas AR, Guarino L and de la Cruz J. (2002). Atlas of wild potatoes. Rome, Italy: International Plant Genetic Resources Institute.
- Hardigan MA, Laimbeer FPE, Newton L, Crisovan E, Hamilton JP, Vaillancourt B, … Buell CR. (2017). Genome diversity of tuber-bearing Solanum uncovers complex evolutionary history and targets of domestication in the cultivated potato. Proc. Natl. Acad. Sci. U. S. A. DOI: 10.1073/pnas.1714380114
- Li Y, Colleoni C, Zhang J, Liang Q, Hu Y, Ruess H, … Huang B. (2018). Genomic analyses yield markers for identifying agronomically important genes in potato. Mol. Plant, 11(3): 473–84. DOI: 10.1016/j.molp.2018.01.009
- Leisner CP, Hamilton JP, Crisovan E, Manrique-Carpintero NC, Marand AP, Newton L, … Buell CR. (2018). Genome sequence of M6, a diploid inbred clone of the high-glycoalkaloid-producing tuber-bearing potato species Solanum chacoense, reveals residual heterozygosity. Plant J., 94(3): 562–70. DOI: 10.1111/tpj.13857
- Bravo-Almonacid F, Rudoy V, Welin B, Segretin ME, Bedogni MC, Stolowicz F, … Mentaberry A. (2012). Field testing, gene flow assessment and pre-commercial studies on transgenic Solanum tuberosum spp. tuberosum (cv. Spunta) selected for PVY resistance in Argentina. Transgenic Res., 21(5): 967–82. DOI: 10.1007/s11248-011-9584-9
- Duan H, Richael C and Rommens CM. (2012). Overexpression of the wild potato eIF4E-1 variant Eva1 elicits Potato virus Y resistance in plants silenced for native eIF4E-1. Transgenic Res., 21(5): 929–38. DOI: 10.1007/s11248-011-9576-9
- Bae J, Halterman D and Jansky S. (2008). Development of a molecular marker associated with Verticillium wilt resistance in diploid interspecific potato hybrids. Mol. Breed., 22(1): 61–9. DOI: 10.1007/s11032-008-9156-8
- Chen L, Guo X, Xie C, He L, Cai X, Tian L, … Liu J. (2013). Nuclear and cytoplasmic genome components of Solanum tuberosum + S. chacoense somatic hybrids and three SSR alleles related to bacterial wilt resistance. Theor. Appl. Genet., 126(7): 1861–72. DOI: 10.1007/s00122-013-2098-5
- Braun SR, Endelman JB, Haynes KG and Jansky SH. (2017). Quantitative trait loci for resistance to common scab and cold-induced sweetening in diploid potato. Plant Genome, 10(3). DOI: 10.3835/plantgenome2016.10.0110
- Jansky S, Douches D and Haynes K. (2017). Transmission of scab resistance to tetraploid potato via unilateral sexual polyploidization. Am. J. Potato Res., 95(3): 272–7. DOI: 10.1007/s12230-017-9628-7
- Almasia NI, Bazzini AA, Hopp HE and Vazquez-Rovere C. (2008). Overexpression of snakin-1 gene enhances resistance to Rhizoctonia solani and Erwinia carotovora in transgenic potato plants. Mol. Plant Pathol., 9(3): 329–38. DOI: 10.1111/j.1364-3703.2008.00469.x
- Faccio P, Vazquez-Rovere C, Hopp E, González G, Décima-Oneto C, Favret E, … Franzone P. (2011). Increased tolerance to wheat powdery mildew by heterologous constitutive expression of the Solanum chacoense. Czech J. Genet. Plant Breed., 47(Special issue): S135–41.
- Mweetwa AM, Hunter D, Poe R, Harich KC, Ginzberg I, Veilleux RE and Tokuhisa JG. (2012). Steroidal glycoalkaloids in Solanum chacoense. Phytochemistry, 75: 32–40. DOI: 10.1016/j.phytochem.2011.12.003
- Lorenzen JH, Balbyshev NF, Lafta AM, Casper H, Tian X and Sagredo B. (2001). Resistant potato selections contain leptine and inhibit development of the Colorado potato beetle (Coleoptera: Chrysomelidae). J. Econ. Entomol., 94(5): 1260–7.
- Manrique-Carpintero NC, Tokuhisa JG, Ginzberg I, Holliday JA and Veilleux RE. (2013). Sequence diversity in coding regions of candidate genes in the glycoalkaloid biosynthetic pathway of wild potato species. G3: Genes, Genomes, Genet., 3(9): 1467–79. DOI: 10.1534/g3.113.007146
- Manrique-Carpintero NC, Tokuhisa JG, Ginzberg I and Veilleux RE. (2014). Allelic variation in genes contributing to glycoalkaloid biosynthesis in a diploid interspecific population of potato. Theor. Appl. Genet., 127(2): 391–405. DOI: 10.1007/s00122-013-2226-2
- Cooper SG, Douches DS and Grafius EJ. (2009). Combining engineered resistance, avidin, and natural resistance derived from Solanum chacoense bitter to control Colorado potato beetle (Coleoptera: Chrysomelidae). J. Econ. Entomol., 102(3): 1270–80.
- Ginzberg I, Thippeswamy M, Fogelman E, Demirel U, Mweetwa AM, Tokuhisa J and Veilleux RE. (2012). Induction of potato steroidal glycoalkaloid biosynthetic pathway by overexpression of cDNA encoding primary metabolism HMG-CoA reductase and squalene synthase. Planta, 235(6): 1341–53. DOI: 10.1007/s00425-011-1578-6
- Molnár I, Besenyei E, Thieme R, Thieme T, Aurori A, Baricz A, … Rakosy-Tican E. (2017). Mismatch repair deficiency increases the transfer of antibiosis and antixenosis properties against Colorado potato beetle in somatic hybrids of Solanum tuberosum + S. chacoense. Pest Manag. Sci., 73(7): 1428–37. DOI: 10.1002/ps.4473
- Crossley MS, Schoville SD, Haagenson DM and Jansky SH. (2018). Plant resistance to Colorado potato beetle (Coleoptera: Chrysomelidae) in diploid F2 families derived from crosses between cultivated and wild potato. J. Econ. Entomol. DOI: 10.1093/jee/toy120
- Boivin N, Morse D and Cappadocia M. (2014). Degradation of S-RNase in compatible pollen tubes of Solanum chacoense inferred by immunogold labeling. J. Cell Sci., 127(Pt 19): 4123–7. DOI: 10.1242/jcs.154823
- Soulard J, Boivin N, Morse D and Cappadocia M. (2014). eEF1A is an S-RNase binding factor in self-incompatible Solanum chacoense. PLoS One, 9(2): e90206. DOI: 10.1371/journal.pone.0090206
- Soulard J, Qin X, Boivin N, Morse D and Cappadocia M. (2013). A new dual-specific incompatibility allele revealed by absence of glycosylation in the conserved C2 site of a Solanum chacoense S-RNase. J. Exp. Bot., 64(7): 1995–2003. DOI: 10.1093/jxb/ert059
- Liu B, Boivin N, Morse D and Cappadocia M. (2012). A time course of GFP expression and mRNA stability in pollen tubes following compatible and incompatible pollinations in Solanum chacoense. Sex. Plant Reprod., 25(3): 205–13. DOI: 10.1007/s00497-012-0192-5
- Aouar L, Chebli Y and Geitmann A. (2010). Morphogenesis of complex plant cell shapes: the mechanical role of crystalline cellulose in growing pollen tubes. Sex. Plant Reprod., 23(1): 15–27. DOI: 10.1007/s00497-009-0110-7
- Parre E and Geitmann A. (2005). Pectin and the role of the physical properties of the cell wall in pollen tube growth of Solanum chacoense. Planta, 220(4): 582–92. DOI: 10.1007/s00425-004-1368-5
- Parre E and Geitmann A. (2005). More than a leak sealant. The mechanical properties of callose in pollen tubes. Plant Physiol., 137(1): 274–86. DOI: 10.1104/pp.104.050773
- Chevalier E, Loubert-Hudon A and Matton DP. (2013). ScRALF3, a secreted RALF-like peptide involved in cell-cell communication between the sporophyte and the female gametophyte in a solanaceous species. Plant J., 73(6): 1019–33. DOI: 10.1111/tpj.12096
- Germain H, Chevalier E, Caron S and Matton DP. (2005). Characterization of five RALF-like genes from Solanum chacoense provides support for a developmental role in plants. Planta, 220(3): 447–54. DOI: 10.1007/s00425-004-1352-0
- Germain H, Gray-Mitsumune M, Houde J, Benhamman R, Sawasaki T, Endo Y and Matton DP. (2013). The Solanum chacoense ovary receptor kinase 11 (ScORK11) undergoes tissue-dependent transcriptional, translational and post-translational regulation. Plant Physiol. Biochem., 70: 261–8. DOI: 10.1016/j.plaphy.2013.05.036
- Germain H, Gray-Mitsumune M, Lafleur E and Matton DP. (2008). ScORK17, a transmembrane receptor-like kinase predominantly expressed in ovules is involved in seed development. Planta, 228(5): 851–62. DOI: 10.1007/s00425-008-0787-0
- Germain H, Houde J, Gray-Mitsumune M, Sawasaki T, Endo Y, Rivoal J and Matton DP. (2007). Characterization of ScORK28, a transmembrane functional protein receptor kinase predominantly expressed in ovaries from the wild potato species Solanum chacoense. FEBS Lett., 581(26): 5137–42. DOI: 10.1016/j.febslet.2007.10.001
- Germain H, Rudd S, Zotti C, Caron S, O’Brien M, Chantha S-C, … Matton DP. (2005). A 6374 unigene set corresponding to low abundance transcripts expressed following fertilization in Solanum chacoense Bitt, and characterization of 30 receptor-like kinases. Plant Mol. Biol., 59(3): 515–32. DOI: 10.1007/s11103-005-0536-8
- Tebbji F, Nantel A and Matton DP. (2010). Transcription profiling of fertilization and early seed development events in a solanaceous species using a 7.7 K cDNA microarray from Solanum chacoense ovules. BMC Plant Biol., 10: 174. DOI: 10.1186/1471-2229-10-174
- Vyetrogon K, Tebbji F, Olson DJH, Ross ARS and Matton DP. (2007). A comparative proteome and phosphoproteome analysis of differentially regulated proteins during fertilization in the self-incompatible species Solanum chacoense Bitt. Proteomics, 7(2): 232–47. DOI: 10.1002/pmic.200600399
- Liu Y, Joly V, Dorion S, Rivoal J and Matton DP. (2015). The plant ovule secretome: a different view toward pollen–pistil interactions. J. Proteome Res., 14(11): 4763–75. DOI: 10.1021/acs.jproteome.5b00618